Gut Microbiota, Succinate, and Chronic Rhinosinusitis
- 1. Department of Otolaryngology and Neck Surgery, Zhujiang Hospital, China
Abstract
Objective: There may be a potential association between gut microbiota and chronic sinusitis. However, there is a lack of population-level studies that provide robust evidence to determine causality.
Methods: Genome-wide association study (GWAS) data for gut microbiota from the Netherlands Microbiome Project and for chronic rhinosinusitis (CRS) from FinnGen were analyzed. The validation group dataset comes from the UK Biobank. MR analyses included the inverse-variance-weighted method, weighted-median estimator, MR-Egger regression, and Weighted mode. Sensitivity analyses involved MR-PRESSO and MR Egger intercept tests, and heterogeneity was assessed using Cochran’s Q statistic. Gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses explored the mechanisms and associated genes.
Results: The IVW method revealed that five groups of gut microbiota causally associated with CRS. Specifically, species Parasutterella excrementihominis (OR=1.10, 95% CI: 1.00-1.21), species Bacteroides faecis (OR=1.07, 95% CI: 1.01-1.13), and the genus Parasutterella (OR=1.09, 95% CI: 1.00-1.19) showed positive associations with CRS, while order Actinomycetales (OR=0.85, 95% CI: 0.74-0.97) and species Bacteroides finegoldii (OR=0.93,95%CI:0.88-0.99) displayed negative correlations. Furthermore, metabolites of inosine monophosphate and urate, the super pathway of arginine polyamine biosynthesis, and glyoxylate TCA were positively correlated with the development of CRS, whereas metabolites of 1,4-dihydroxy-2-naphthalate, lysine and peptidoglycan, and the super pathway of Lipid A biosynthesis exhibited inverse causal relationships with CRS. UK Biobank data confirm the causal relationship of the tricarboxylic acid cycle in chronic rhinosinusitis and gut microbiota. Reverse MR analysis indicated CRS could inhibit gut-associated reverse TCA cycle. GO and KEGG analysis identified SUCNR1-related genes as key in chronic sinusitis and intestinal microbiota.
Conclusions: There is a mutual causal relationship between gut microbiota and chronic sinusitis, suggesting that modulating gut microbiota could improve CRS. Succinic acid of intestinal origin may be a mediator of intestinal flora and chronic sinusitis. However, this f inding still needs our further is experimental verification.
Keywords
• Gut microbiota
• Chronic sinusitis
• Mendelian analysis
• SUCNR1
• Nasal polyp
Citation
Hu X, Li J, Chen F, Chen M, Liang T, et al. (2026) Gut Microbiota, Succinate, and Chronic Rhinosinusitis. J Ear Nose Throat Disord 10(1): 1059.
ABBREVIATIONS
CRS: Chronic Rhinosinusitis; GEO: Gene Expression Omnibus; GWAS: Genome wide association study; MR: Mendelian Randomization; GO: Gene Ontology; KEGG: Kyoto Encyclopedia of Genes and Genomes enrichment; IVW: Inverse-variance weighted; OR: Odds ratio; CI:Confidence Interval; UK: United Kingdom; ECRSwNP : Eosinophilic chronic rhinosinusitis with nasal polyps; NECRSwNP: Non-eosinophilic chronic rhinosinusitis with nasal polyps; GWAS: Genome wide association study; CRSwNP: Chronic rhinosinusitis with nasal polyps; SNPs: Single nucleotide Polymorphisms; RCTs: Randomized Controlled trials; IVs: Instrumental Variables; SUCNR1: Succinate Receptor 1; TCA: Tricarboxylic acid; DSS: Dextran Sulfate Sodium; LPS: Lipopolysaccharide; VK2: Vitamin K2; SCC: Solitary Chemosensory Cell; scRNA-seq: Single cell Ribonucleic acid sequencing.
INTRODUCTION
The human gastrointestinal tract harbors numerous microorganisms, which play pivotal roles in regulating the body’s physiological functions including endocrine, neurological, and immune functions [1]. Dysbiosis of intestinal microbiota, intestinal barrier dysfunction,increased intestinal permeability, and leakage of toxic bacterial metabolites can promote the occurrence of systemic inflammatory diseases [2,3]. Previous observational studies have shown a correlation between gut microbiota and respiratory tract infections [4,5]. Furthermore, research has demonstrated the effectiveness of oral probiotics, such as Bifidobacterium and Lactobacillus, in reducing sinus infectictious [6,7]. Dysbiosis of intestinal microbiota has been observed in patients with eosinophilic chronic rhinosinusitis with nasal polyps (ECRSwNP) and non-eosinophilic chronic rhinosinusitis with nasal polyps (NECRSwNP). In both types of chronic rhinosinusitis with nasal polyps (CRSwNP), there was a significant decrease in the abundance of Actinomycetes and its predominant genus, Bifidobacteria, with ECRSwNP showing a more pronounced decrease [7]. However, establishing a causal relationship between gut microbiota and chronic rhinosinusitis in clinical trials is challenging due to the influence of underlying factors. MR offers a method to explore causal relationships between exposures and outcomes using genetic variation as instrumental variables [8]. By employing single nucleotide polymorphisms (SNPs) as instrumental variables, we can assess the impact of microbiota on infection while minimizing confounding effects, unlike artificially designed or traditional randomized controlled trials (RCTs [9]. In this study, we investigate the causal relationship between gut microbiota and CRS through a comprehensive bidirectional MR analysis derived from the IEU Open GWAS project. Through this analysis, we aim to elucidate the role of gut microbiota and its associated metabolic pathways in the development of chronic rhinosinusitis, thereby contributing to the exploration of novel therapeutic strategies such as probiotic therapy and dietary modifications.
MATERIALS AND METHODS
Gut microbiota
GWAS data on gut microbiota were obtained from the Dutch Microbiome Project. This study encompassed 7,738participants, analyzing 207taxa and 205pathways representing microbial composition and function. Full GWAS summary statistics are accessible in the NHGRI-EBI GWAS Catalog (https://www.ebi.ac.uk/gwas/).
Chronic rhinosinusitis
Summary-level statistics were acquired from the FinnGen project, comprising 176,373 participants, including 8,524CRS cases and 167,849 controls. CRS is defined as a chronic inflammatory process affecting the mucosa of any sinus cavity. Validation group data comprising 2888patients with chronic rhinosinusitis and 460045 controls from UK Biobank (GWAS id: UKB-b-69).
Mendelian Randomization analysis
All data analysis was performed in the R environment (version 4.3.1) and in the R package: two-sample MR [10], MR-Egger [11], and MR-PRESSO [12]. Initially, SNPs significantly associated with gut microbiota and microbial pathways were selected as instrumental variables (IVs), with a significance threshold set at 5×10^-8 [13]. Due to a reduced number of SNPs meeting this criterion for the remaining intestinal microbiota, the threshold was adjusted to 1×10^-5, though the criterion for reverse MR analysis remained stringent at 5×10^-8. Subsequently, we excluded SNPs in linkage disequilibrium using a linkage disequilibrium parameter of 0.01 and a genetic distance of 10,000 kb [14]. We then employed the MR-PRESSO regression test and MR Egger intercept to assess potential horizontal pleiotropic effects. The remaining SNPs, after the removal of pleiotropic SNPs, were used for subsequent MR analysis. Four methods (inverse variance weighted, MR Egger, weighted median, and weighted mode) were utilized to assess the genetic prediction-specific association between gut microbiota and chronic rhinosinusitis. IV [15], method served as the primary fixed-effect meta-analysis, with the remaining three methods used for secondary validation to enhance result reliability. A significance level of IVW-P< 0.05 was considered an initial positive result. Finally, to investigate whether CRS causally influenced gut flora, we conducted a reverse MR analysis, treating CRS as exposure data and gut flora as outcome data. Cochran’s Q statistic was employed to assess heterogeneity in IVW meta-analyses, with P< 0.05 indicating heterogeneity in SNP effect estimates. A non-significant result in both MR PRESSO and MR Egger intercept tests (P > 0.05) indicated that the analysis was unaffected by horizontal plei [16-25].
DISCUSSION
There is growing evidence of a potential causal relationship between gut microbiota and CRS. This study not only found that the gut microbiota has an effect on CRS, but CRS also interferes with the gut microbiota. Among gut microbes, Parasutterella and Bacteroides contribute to CRS. The genus of Parasutterella has been defined as a core component of the human and mouse gut microbiota, and has been correlated with various health outcomes [26]. It has been shown that Parasutterella produces succinate as a fermentation end product. Succinate serves as one of the key intermediate metabolites produced by intestinal flora [27,28], and the ability of Parasutterella to produce succinate suggests a potential way for Parasutterella to support interspecies metabolic interactions within the intestinal ecosystem [29].
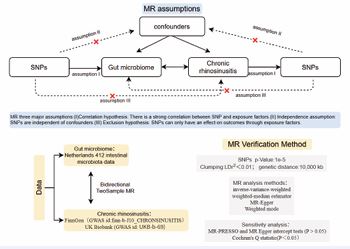
Figure 1 A bidirectional two-sample MR model was used to evaluate the causal relationships between exposure and outcome
In the host, succinate, an important pro-inflammatory signal through IL-1β, has been shown to be a key mediator of the macrophage response to lipopolysaccharide. And both recruit eosinophils and inflammatory factors, promoting type 2 inflammation and exacerbating CR [30,31]. Succinate production in feces has been positively correlated with the relative abundance of Bacteroides and Parasutterella in a dextran sulfate sodium (DSS) induced colitis mouse mode [32]. It is possible that the same pathway exists for the effects of both gut microbes on CRS. MR analysis also revealed Actinomycetes as a protective factor for CRS. Actinomycetes are involved in tryptophan metabolism in the gut [33]. In observational studies, the abundance of Actinobacteria phylum was significantly higher in elderly patients, which was positively correlated with the biosynthesis of several aromatic amino acid [34]. Airway microbiomics and serum metabolomics show a correlation between Actinomycetes and tryptophan metabolism [35]. The tryptophan metabolite indole acrylic acid promotes the restoration of intestinal epithelial barrier function and attenuates the inflammatory response [36,37]. Bacteroides finegoldii, as a classical chondroitin sulphate degrader, may inhibit the development of CRS by producing large amounts of short chain fatty acids, such as acetate, propionate and butyrate, as a result of the fermentation of chondroitin sulphate in the human intestine [38].
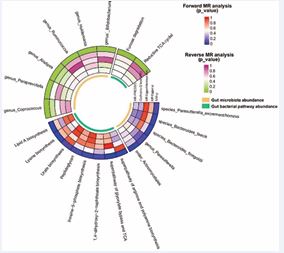
Figure 2 Circumferential heat map of the correlation between intestinal flora and chronic sinusitis. CRS: chronic Rhinosinusitis. Forward MR: Intestinal flora as exposure, Reverse MR: CRS as exposure. MR-PRESSO, Mendelian Randomization Pleiotropy Residual Sum and Outlier. IVW-p, inverse-variance weighted method, p?0.05was positive. MR Egger-p: Mendelian Randomization egger, P?0.05was positive. Cochran’s Q test (IVW): Mendelian Randomization Heterogeneity Examination, P > 0.05, indicating no heterogeneity. MR egger intercept test: P>0.05, indicating no horizontal pleiotropy.
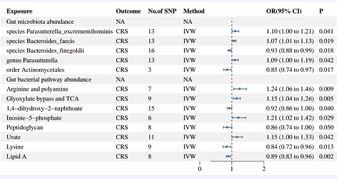
Figure 3 Forest plot of positive MR results. Gut flora as exposure data and CRS as outcome data.
Positive promotion of the abundance of intestinal metabolites in CRS includes urate and inosine monophosphate. Metabolomics and observational studies have found that NECRSwNP was characterized by uric acid accumulation. Elevated uric acid levels are positively correlated with mucosal neutrophil numbers and mRNA levels of IFN-γ, IL-17A, IL-1β, and IL-8 [39]. In addition, there are pathways for inosine monophosphate and uric acid conversion [40]. In the future, uric acid may be an important marker for predicting CRS [41]. Another important result of the analysis was the correlation of amino acid metabolites, further reflecting the potential role of amino acids in CRS [42]. Similarly, disorders of amino acid metabolism are common in asthma and intestinal inflammation. It is important to note that lipid A is inversely related to the occurrence of CRS. Lipid A is a glucosamine disaccharide that is a major component of lipopolysaccharide (LPS) and is a gram-negative extra adventitial leaflet [43]. As an antibiotic target and immunotherapy molecule, lipid A deserves our attention in the future research on CRS. 1,4-dihydroxy-2-naphthalate is an intermediate in the biosynthesis of naphthoketone (vitamin K2) by genus Escherichia. Existing studies have found VK2 to be beneficial in type 2 diabetes-related inflammation by increasing the concentration of anti inflammatory n-3 polyunsaturated fatty acids [44]. Others have reported that VK2 is effective in mitigating acute lung injury by inhibiting lipopolysaccharide induced inflammation, apoptosis, iron death and elastin degradation [45]. We suspect that the protective effect of 1,4-dihydroxy-2-naphthalate against CRS is generated through the anti-inflammatory effect of VK2.Inverse MR analysis showed that CRS was inversely correlated with six species of gut bacteria. Previous controlled experiments likewise yielded similar results.
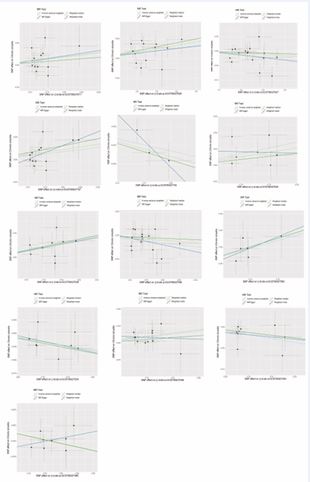
Figure 4 Scatter plots of all positive outcomes with CRS as exposure and gut microbiota as outcome ter plots of all positive outcomes with CRS as exposure and gut microbiota as outcome.
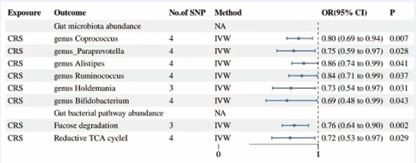
Figure 5 Forest plot results for CRS as exposure and gut flora and its metabolites as outcome.
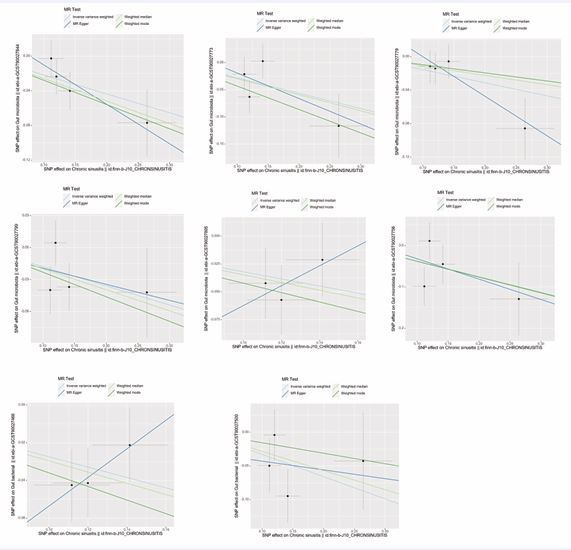
Figure 6 Scatter plots of all positive outcomes with gut microbiota as exposure and CRS as outcome.

Figure 7 Boxplot of differential expression of SUCNR1 in nasal polyp tissue versus normal tissue. SUCNR1 gene was significantly up-regulated in nasal polyp tissues (P<0.05). Bubble plot of the results of signaling pathway enrichment analysis in the differentially expressed SUCNR1 gene (KEGG). Bubble maps of the results of functional enrichment analysis in differentially expressed genes, including biological processes, cellular components, and molecular functions.
Using bidirectional MR analysis, we were pleasantly surprised to find a factor that correlated both CRS and gut microbiota, which is the tricarboxylic acid cycle (TCA). Among these, the superpathways of glyoxylate bypass and TCA(OR=1.15) promote CRS, and the reductive TCA cycle I(OR=0.72) pathway inhibits CRS. The TCA cycle is a catabolic pathway that generates energy and reducing power as well as precursors for biosynthesis [46,47]. We selected succinate, a common intermediate of both metabolic pathways, as the main research site. Studies have found that succinate is mainly produced by the intestines. Succinate binds to the SUCNR1 receptor in the human body and produces important inflammatory modulatory effects [48,49]. In nasal polyps, we found higher SUCNR1 expression, which was also found in previous studies. The solitary chemosensory cell (SCC) is the predominant source of IL-25 in the sinonasal epithelium [50]. SCC expresses the succinate receptor SUCNR1 and regulates the type 2 inflammatory response to intestinal helminths and protozoan infections. SCC is the primary epithelial source of interleukin-25 and an important source of cysteinyl leukotrienes in the airways and are thought to be upstream agents for CRS and asthma type 2 inflammation. Studied scRNAseq analysis showed that approximately 10% of the SCC/ionocyte clusters expressed SUCNR1 and a small population of immune cells. Various evidence suggests that succinate plays an important role in the pathogenesis of CRS.
Table 1: Summary of causality between Gut flora and CRS.
|
Exposure |
Outcome |
No. of SNP |
MR results |
|
|
MR Egger Intercept test |
Cochrans’Q test (IVW) |
MR- PRESSO |
|
Gut microbiota abundance species Parasutterella excrementihominis species Bacteroides faecis species Bacteroides finegoldii genus Parasutterella order Actinomycetales Gut bacterial pathway abundance Arginine and polyamine Glyoxylate bypass and TCA 1,4-dihydroxy-2-naphthoate Inosine-5-phosphate Peptidoglycan Urate Lysine Lipid A |
|
|
Method |
OR(95%CI) |
P-value |
|
|
|
|
CRS |
13 |
IVW |
1.104(1.004,1.213) |
0.0401 |
|
|
|
|
|
|
MR Egger |
1.128(0.638,1.992) |
0.687 |
|
|
|
||
|
|
|
|
0.942 |
0.336 |
0.377 |
|||
|
Weighted median |
1.097 (0.966,1.247) |
0.154 |
|
|
|
|||
|
Weighted mode |
1.062 (0.869,1.298) |
0.569 |
|
|
|
|||
|
13 |
IVW |
1.069(1.011,1.129) |
0.019 |
|
|
|
||
|
|
|
|
0.943 |
0.205 |
0.238 |
|||
|
MR Egger |
1.060(0.842,1.334) |
0.631 |
|
|
|
|||
|
Weighted median |
1.088(1.014,1.167) |
0.019 |
|
|
|
|||
|
Weighted mode |
1.104(0.985,1.239) |
0.116 |
|
|
|
|||
|
16 |
IVW |
0.934(0.883,0.988) |
0.018 |
0.838 |
0.307 |
0.317 |
||
|
MR Egger |
0.914(0.741,1.128) |
0.418 |
|
|
|
|||
|
Weighted median |
0.963(0.891,1.034) |
0.334 |
|
|
|
|||
|
Weighted mode |
0.978(0.866,1.105) |
0.726 |
|
|
|
|||
|
13 |
IVW |
1.094(1.003,1.193) |
0.042 |
0.282 |
0.524 |
0.553 |
||
|
MR Egger |
1.355(0.926,1.983) |
0.146 |
|
|
|
|||
|
Weighted median |
1.120(0.996,1.260) |
0.058 |
|
|
|
|||
|
Weighted mode |
1.169(0.964,1.417) |
0.139 |
|
|
|
|||
|
3 |
IVW |
0.850(0.744,0.971) |
0.017 |
0.458 |
0.425 |
0.213 |
||
|
MR Egger |
0.489(0.188,1.274 |
0.381 |
|
|
|
|||
|
Weighted median |
0.819(0.690,0.972) |
0.022 |
|
|
|
|||
|
Weighted mode |
0.800(0.643,0.994) |
0.182 |
|
|
|
|||
|
CRS |
7 |
IVW |
1.240(1.055,1.457) |
0.009 |
0.463 |
0.308 |
0.37 |
|
|
|
MR Egger |
0.973(0.523,1.811) |
0.936 |
|
|
|
||
|
Weighted median |
1.175(0.961,1.437) |
0.116 |
|
|
|
|||
|
Weighted mode |
1.117(0.851,1.466) |
0.456 |
|
|
|
|||
|
9 |
IVW |
1.146(1.041,1.262) |
0.005 |
0.982 |
0.683 |
0.698 |
||
|
MR Egger |
1.152(0.764,1.735) |
0.521 |
|
|
|
|||
|
Weighted median |
1.159(1.016,1.323) |
0.028 |
|
|
|
|||
|
|
Weighted mode |
1.164(0.951,1.426) |
0.179 |
|
|
|
||
|
15 |
IVW |
0.924(0.857,0.996) |
0.04 |
0.662 |
0.631 |
0.647 |
||
|
MR Egger |
0.865(0.642,1.165) |
0.357 |
|
|
|
|||
|
Weighted median |
0.951(0.856,1.057) |
0.349 |
|
|
|
|||
|
Weighted mode |
0.971(0.828,1.138) |
0.723 |
|
|
|
|||
|
6 |
IVW |
1.205(1.019,1.425) |
0.029 |
0.826 |
0.937 |
0.946 |
||
|
MR Egger |
1.312(0.634,2.716) |
0.505 |
|
|
|
|||
|
Weighted median |
1.228(0.993,1.518) |
0.059 |
|
|
|
|||
|
Weighted mode |
1.227(1.936,1.608) |
0.199 |
|
|
|
|||
|
8 |
IVW |
0.859(0.737,1.000) |
0.049 |
0.837 |
0.695 |
0.729 |
||
|
MR Egger |
0.816(0.500,1.331) |
0.446 |
|
|
|
|||
|
Weighted median |
0.852(0.701,1.036) |
0.108 |
|
|
|
|||
|
Weighted mode |
0.833(0.628,1.105) |
0.245 |
|
|
|
|||
|
11 |
IVW |
1.155(1.005,1.327) |
0.042 |
0.583 |
0.948 |
0.953 |
||
|
MR Egger |
0.954(0.486,1.871) |
0.894 |
|
|
|
|||
|
Weighted median |
1.172(0.972,1.412) |
0.096 |
|
|
|
|||
|
Weighted mode |
1.091(0.816,1.460) |
0.569 |
|
|
|
|||
|
9 |
IVW |
0.835(0.725,0,963) |
0.013 |
0.757 |
0.438 |
0.495 |
||
|
MR Egger |
0.899(0.561,1.438) |
0.669 |
|
|
|
|||
|
Weighted median |
0.879(0.728,1.060) |
0.177 |
|
|
|
|||
|
Weighted mode |
0.880(0.686,1.130) |
0.347 |
|
|
|
|||
|
8 |
IVW |
0.892(0.829,0.959) |
0.002 |
0.319 |
0.871 |
0.889 |
||
|
MR Egger |
1.106(0.744,1.644) |
0.635 |
|
|
|
|||
|
Weighted median |
0.893(0.813,0.982) |
0.019 |
|
|
|
|||
|
Weighted mode |
0.892(0.779,1.021) |
0.141 |
|
|
|
This study has the following advantages. First, we explored the bidirectional causal association between gut microbiota and CRS by using a bidirectional MR design that was less susceptible to confounders, reverse causality, and non-differential exposures. Second, we conducted sensitivity analyses such as MR-Egger, weighted median, and MR-PRESSO to confirm the consistency and reliability of our results. Third, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses helped identify a possible link between succinic acid, a gut microbiota metabolite, and CRS. However, our study has some limitations. The research sample consisted exclusively of individuals from the European population, limiting the generalizability of our findings to other demographics. Additionally, further experimental studies are required to validate and elaborate on the proposed mechanisms.
CONCLUSION
This study shows that there is a causal relationship between gut microbiota and chronic sinusitis. Succinic acid, an intermediate product of the TCA cycle associated with intestinal metabolism, plays an important role. Succinate may be an important target for the diagnosis and treatment of CRS. In addition, it may be possible to improve CRS by dietary regulation of intestinal succinate concentrations.
Ethics approval
The authors declare that there is no requirement for ethical approval for this submission. This is because the data used in the study are all from public databases.
ACKNOWLEDGEMENTS
We thank the participants and researchers at FinnGen, UK Biobank and GWAS Catalog for providing publicly available GWAS summary data for this study. We also thank TCGA and the GEO database for their platforms and the contributors who uploaded significant datasets.
AUTHOR CONTRIBUTIONS
XY H, JL and FC conceptualized the research and analysed the dataset. TF L, YH, and XY H wrote the first draft of the manuscript. data analyses:XY H, JL, MC, SJ C, CS Y interpreted the results, revised the paper. FC, CS Y and MY provided critical intellectual content.
FUNDING
This work was sponsored by the Natural Science Foundation of Guangdong Province (2023A1515030291), Science and Technology Program of Guangzhou, China (202201020017) and Dean’s Fund Project [rzjj2024ms08]
AVAILABILITY OF DATA AND MATERIALS
The UK Biobank is an open access resource. Researchers can apply to use the UK Biobank dataset by registering and applying at [http://ukbiobank.ac.uk/register-apply/]. Full GWAS summary statistics are accessible in the NHGRI-EBI GWAS Catalog [https://www.ebi.ac.uk/gwas/].
REFERENCES
- Adak A, Khan MR. An insight into gut microbiota and its functionalities.Cell Mol Life Sci. 2019; 76: 473-493.
- Mou Y, Du Y, Zhou L. Gut Microbiota Interact with the Brain Through Systemic Chronic Inflammation: Implications. Front Immunol. 2022; 13: 6279-6288.
- Malesza IJ, Malesza M, Walkowiak J. High-Fat, Western-Style Diet, Systemic Inflammation, and Gut Microbiota: A Narrative Review. Cells. 2021; 10: 3164.
- Budden KF, Gellatly SL, Wood DLA. Emerging pathogenic links between microbiota and the gut-lung axis. Nat Rev Microbiol. 2017; 15: 55-63.
- de Steenhuijsen Piters WAA, Binkowska J, Bogaert D. Early Life Microbiota and Respiratory Tract Infections. Cell Host Microbe. 2020; 28: 223-232.
- Cope EK, Lynch SV. Novel microbiome-based therapeutics for chronic rhinosinusitis. Curr Allergy Asthma Rep. 2015; 15: 30.
- Abbas EE, Li C, Xie A. Distinct Clinical Pathology and Microbiota in Chronic Rhinosinusitis with Nasal Polyps Endotypes. Laryngoscope. 2021; 131: E34-E44.
- Xu J, Li M, Gao Y. Using Mendelian randomization as the cornerstone for causal inference in epidemiology. Environ Sci Pollut Res Int. 2022; 29: 5827-5839.
- Ference BA, Holmes MV, Smith GD. Using Mendelian Randomization to Improve the Design of Randomized Trials. Cold Spring Harb Perspect Med. 2021; 11: a040645.
- Hemani G, Zheng J, Elsworth B. The MR-Base platform supports systematic causal inference across the human phenome. Elife. 2018; 7: e34408.
- Burgess S, Thompson SG. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur J Epidemiol. 2017; 32: 377-389.
- Verbanck M, Chen CY, Neale B, Do R. Detection of widespread horizontal pleiotropy in causal relationships. Nat Genet. 2018; 50: 693-698.
- Sanna S, van Zuydam NR, Mahajan A. Causal relationships among the gut microbiome. Nat Genet. 2019; 51: 600-605.
- Kamat MA, Blackshaw JA, Young R. PhenoScanner V2: an expanded tool for searching human genotype-phenotype associations. Bioinformatics. 2019; 35: 4851-4853.
- Burgess S, Butterworth A, Thompson SG. Mendelian randomization analysis. Genet Epidemiol. 2013; 37: 658-665.
- Prediction and analysis of essential genes using the enrichments of gene ontology and KEGG pathways.
- MM, APK, Am B, MS, MG, AS. Alteration of indicator gut microbiota in patients with chronic sinusitis. Immunity, Inflammation and Disease. 2023; 11: 997-999.
- Wan Y, Yuan J, Li J. Overweight and underweight status are linked to specific gut microbiota. Clin Nutr. 2020; 39: 3189-3198.
- Strobel KM, Del Vecchio G, Devaskar SU, Calkins KL. Gut microbes and circulating cytokines in preterm infants. J Nutr. 2023; 153: 120-130.
- Fan L, Zhu X, Sun S. Ca: Mg ratio, medium-chain fatty acids, and the gut microbiome. Clin Nutr. 2022; 41: 2490-2499.
- Nicolau SE, Lewis K. The Role of Integration Host Factor in Escherichia coli Persister Formation. mBio. 2022; 13: e03420-421.
- Tong W, Hannou SA, Wang Y. The intestine is a major contributor to circulating succinate in mice. FASEB J. 2022; 36: e225-246.
- E M, La O. Succinate: a metabolic signal in inflammation. Trends Cell Biol. 2014; 24: 24-25.
- Yh W, Zz Y, Sd L. Gut microbiota-derived succinate aggravates acute lung injury following intestinal ischaemia/reperfusion in mice. Eur Respir J. 2023; 61
- Gilissen J, Jouret F, Pirotte B, Hanson J. Insight into SUCNR1 (GPR91) structure and function. Pharmacol Ther. 2016; 159: 56-65.
- Willing BP, Dicksved J, Halfvarson J. Gastrointestinal microbial profiles vary with inflammatory bowel disease phenotypes. Gastroenterology. 2010; 139: 1844-1854.
- De Vadder F, Kovatcheva-Datchary P, Zitoun C. Microbiota- Produced Succinate Improves Glucose Homeostasis via Intestinal Gluconeogenesis. Cell Metab. 2016; 24: 151-157.
- Macias-Ceja DC, Ortiz-Masiá D, Salvador P. Succinate receptor mediates intestinal inflammation and fibrosis. Mucosal Immunol. 2019; 12: 178-187.
- Fischbach MA, Sonnenburg JL. Eating for two: how metabolism establishes interspecies interactions in the gut. Cell Host Microbe. 2011; 10: 336-347.
- Wang W, Xu Y, Wang L. Single-cell profiling identifies mechanisms of inflammatory heterogeneity in chronic rhinosinusitis. Nat Immunol. 2022; 23: 1484-1494.
- Wang Z, Lin D, Zhao Y, Liu H, Yang T, Li A. MiR-214 Expression Is Elevated in Chronic Rhinosinusitis Mucosa. Am J Rhinol Allergy. 2023; 37: 391-401.
- Osaka T, Moriyama E, Arai S. Meta-Analysis of Fecal Microbiota and Metabolites in Experimental Colitic Mice. Nutrients. 2017; 9: 1329.
- Fan J, Zhou Y, Meng R. Cross-talks between gut microbiota and tobacco smoking: a two-sample Mendelian randomization study. BMC Med. 2023; 21: 16.
- Chen F, Gao W, Yu C. Age-Associated Changes of Nasal Bacterial Microbiome in Patients with Chronic Rhinosinusitis. Front Cell Infect Microbiol. 2022; 12: 788141.
- Yuan Y, Wang C, Wang G. Airway Microbiome and Serum Metabolomics Identify Differential Candidate Biomarkers in Allergic Rhinitis. Front Immunol. 2021; 12: 1077-1136.
- Wlodarska M, Luo C, Kolde R. Indoleacrylic acid produced by commensal Peptostreptococcus species suppresses inflammation. Cell Host Microbe. 2017; 221-225.
- Grifka-Walk HM, Jenkins BR, Kominsky DJ. Amino Acid Trp: The Far Out Impacts of Host and Commensal Tryptophan Metabolism. Front Immunol. 2021; 12: 653-708.
- Wang Y, Ma M, Dai W, Shang Q, Yu G. Bacteroides salyersiae is a potent chondroitin sulfate-degrading species in the human gut microbiota. Microbiome. 2024; 12: 141.
- Li JX, Wang ZZ, Zhai GT. Untargeted metabolomic profiling identifies disease-specific and outcome-related signatures in chronic rhinosinusitis. J Allergy Clin Immunol. 2022; 150: 727-735.
- Andres-Hernando A, Cicerchi C, Kuwabara M, et al. Umami-induced obesity and metabolic syndrome is mediated by nucleotide degradation and uric acid generation. Nat Metab. 2021; 3: 1189-1201.
- Xie S, Zhang C, Xie Z, Zhang J, Zhang H, Jiang W. Serum metabolomics identifies uric acid as a possible novel biomarker for predicting recurrence of chronic rhinosinusitis with nasal polyps. Rhinology. 2023; 61: 541-551.
- He F, Wu C, Li P. Functions and Signaling Pathways of Amino Acids in Intestinal Inflammation. Biomed Res Int. 2018; 2018: 9171895.
- Garcia-Vello P, Di Lorenzo F, Zucchetta D, Zamyatina A, De Castro C, Molinaro A. Lipopolysaccharide lipid A: A promising molecule for new immunity-based therapies and antibiotics. Pharmacol Ther. 2022; 230: 107970
- Ko?akowski A, Kurzyna PF, Bzd?ga W. Influence of vitamin K2 on lipid precursors of inflammation and fatty acid pathway activities in HepG2 cells. Eur J Cell Biol. 2021; 100: 7-8.
- Wang Y, Yang W, Liu L. Vitamin K2 (MK-7) attenuates LPS-induced acute lung injury. PLoS One. 2023; 18: e40290.
- Martínez-Reyes I, Chandel NS. Mitochondrial TCA cycle metabolites control physiology and disease. Nat Commun. 2020; 11: 102.
- Casadio M. Nuclear TCA cycle reactions. Nat Cell Biol. 2022; 24: 1448.
- Trauelsen M, Hiron TK, Lin D. Extracellular succinate hyperpolarizes M2 macrophages. Cell Rep. 2021; 35: 109246.
- Perniss A, Boonen B, Tonack S. A succinate/SUCNR1-brush cell defense program in the tracheal epithelium. Sci Adv. 2023; 9: eadg884902.
- Kohanski MA, Workman AD, Patel NN. Solitary chemosensory cells are a primary epithelial source of IL-25 in patients with chronic rhinosinusitis with nasal polyps. J Allergy Clin Immunol. 2018; 142: 460-470.









































































































































{kind=link}