Comparison of Effects of Chenodeoxycholic Acid and Cholic acid on Diabetic Nephropathy in Kkay Mice
- 1. Department of Internal Medicine, Dokkyo Medical University, Japan
Abstract
Aims: The main purpose of the present study was to investigate the effects of chenodeoxycholic acid (CDCA) and cholic acid (CA) on diabetic nephropathy in KKAy mice, an animal model of type 2 diabetes.
Methods: Ten-week-old male KK-Ay/Ta Jcl (KKAy) mice were randomly divided into 3 groups composed of 8 mice each; CDCA, CA and no drugs. C57BL/6J mice were used as control. Parameters were evaluated at 22 weeks of age.
Results: CA significantly decreased casual glucose levels and increased total bile acid. There were no significant differences in systolic and diastolic blood pressure among the KKAy control, CDCA and CA groups. Both CDCA and CA significantly decreased UAE, and mRNA expression of MCP-1 and fibronectin in the kidney, which corresponded to improved renal histological findings.
Conclusion: These results suggest that CA, but not CDCA improves, glycemic control as reflected by casual BG levels, but that CDCA and CA similarly inhibit the progression of diabetic nephropathy in KKAy mice.
Citation
Takebayashi K, Inukai T (2014) Comparison of Effects of Chenodeoxycholic Acid and Cholic acid on Diabetic Nephropathy in Kkay Mice. J Endocrinol Diabetes Obes 2(3): 1053.
INTRODUCTION
It was recently reported that cholic acid (CA), a representative primary bile acid in humans [1], improves glycemic control in a mouse model of type 2 diabetes [2,3]. Although the mechanisms of the glucose-lowering effects of bile acid have not been fully elucidated, inhibition of gluconeogenesis in the liver via the farneoid X receptor (FXR)-small heterodimer partner (SHP)- phosphoenolpyruvate carboxykinase (PEPCK) pathway [3], increased glucose metabolism by regulating energy homeostasis via activation of the G protein-coupled receptor 5 (TGR5)- cyclic adenosine monophosphate (cAMP)-type 2 iodothyronine deiodinase (D2) pathway in brown adipose tissue or skeletal muscle [4,5], and increased secretion of glucagon like peptide-1 (GLP-1), which promotes insulin secretion in pancreatic b-cells, in L cells in the intestine via TGR5 [6-8] may be partially involved.
FXR is expressed in both the liver and the kidney [9-11], and CA reportedly inhibits the progression of diabetic nephropathy by mechanisms via FXR in the kidney independently of its glucose-lowering effects [2]. It is known that chenodeoxy cholic acid (CDCA), another primary bile acid in humans, has a stronger FXR-agonist effect than CA [1]; therefore, CDCA may have more beneficial effects on inhibition of diabetic nephropathy than CA. However, the effects of CDCA on diabetic nephropathy and glycemic control have not yet been fully elucidated. In the present study, we compared the effects of CDCA and CA on the progression of diabetic nephropathy in KKAy mice, a mouse model of type 2 diabetes.
MATERIALS AND METHODS
Animals and study design
Animal studies were conducted according to the “Guiding Principles for the Care and Use of Laboratories Animals” of the Japanese Pharmacological Society. Ten-week-old male KK-Ay/Ta Jcl (KKAy) mice were purchased from CLEA Japan (Tokyo, Japan). KKAy mice were randomly divided into 3 groups composed of 8 mice each; no drug treatment (KKAy control), and treatment with CDCA or CA. C57BL/6J mice were used as control (n =8). Mice were individually housed in plastic cages with free access to food (CLEA rodent diet CE-2®: 343.1 kcal/100 g, CLEA Japan) and water throughout the 12-week experimental periods. Drugs were administered by mixing into feed (0.5% (wt/wt) for both CDCA and CA)). CDCA and CA powder was purchased from Wako Pure Chemical Industries, Ltd. (Osaka, Japan).
At baseline (age, 10 weeks), no significant differences were observed in casual BG, SBP and DBP among the control C57BL/6J mice, and the 3 KKAy groups. Significantly higher BW was observed in the 3 KKAy groups when compared with the C57BL/6J mice, while no significant differences in BW were evident among the 3 KKAy groups. All mice were maintained in the same room under conventional conditions with a regular 12-h light/dark cycle and temperature of 24°C.
Based on data provided from CLEA Japan Tokyo, Japan), most mice do not show proteinuria at 5 weeks of age, but 1+ to 2+ proteinuria is seen in all KKAy mice at 10 weeks of age, while 2+ to 3+ proteinuria is observed in all mice at 18 weeks of age. We selected 10-week-old mice as the initiation point, as this is considered to be the stage of mild or moderate diabetic nephropathy.
Measurement of BW and BP, and blood and urine tests
Measurement of BW, BP and casual BG was performed at 10, 14, 18 and 22 weeks of age. BP was measured in the morning by a non-invasive tail cuff using apparatus for measuring blood pressure for mice (BP-98A®, Softron Co., Ltd, Tokyo) after mice were pre-warmed for 15 min at 37°C using Animal Blanket Controller ATB-1100® (Nihon Kohden Co., Ltd., Tokyo, Japan). A total of 3 measurements were performed in each mouse. BG was measured in tail capillary vessel blood using a blood glucose meter (Medisafe-mini® GR-102, Terumo Co., Ltd, Tokyo, Japan).
Blood for measurement of circulating lipids and total bile acids (TBA) was obtained from the heart when mice were sacrificed at 22 weeks of age. Serum TG, HDL-C, LDL-C and TBA were measured using N-Assay-L TG-H Nittobo® (Nittobo), MetaboLead HDL-C® (Kyowa Medex Co., Ltd., Tokyo, Japan), MetaboLead LDL-C® (Kyowa Medix), and Aqua-auto Kainos TBA® (KAINOS Laboratories Inc., Tokyo, Japan) respectively.
Urine samples were collected at 22 weeks of age using cages. Measurement of urinary albumin and creatinine was performed using an Albumin Mouse Urine ELISA kit, Albuwell M® (Exocell Inc., Philadelphia, PA), and a Creatinine Companion Kit® (Exocell), respectively.
Light microscopy and immunohistochemical staining
Renal cortex specimens (3-μm sections) were prepared for light microscopy by periodic acid-Schiff (PAS) staining. With regard to immunochemistry, 3-μm sections were prepared using a cryostat. Sections were fixed in cold acetone at -20°C for 10 min. Cryosections were incubated overnight with antibodies against MCP-1 (1:100 dilution) (MCP-1 (FL-148), sc-28879®, Santa Cruz Biotechnology Inc., CA, USA) and p47phox (1:100 dilution) (p47-phox(H-195), sc-14015®, Santa Cruz Biotechnology) after blocking with blocking solution. Slides were examined using an Olympus AX80® (Olympus Co., Ltd., Tokyo, Japan).
In order to quantitatively evaluate the extent of glomerular sclerosis, 3 glomeruli were randomly selected for each mouse in all groups. The PAS-positive area in the glomerular mesangium was defined as the extent of extracellular mesangium matrix area (ECM), and ECM/whole glomerular area (WGA)(%) was used as a surrogate marker of the extent of glomerular sclerosis. The mean of all glomeruli selected in each group (i.e., n×3) was defined as ECM/WGA.
MCP-1 and p47-phox protein production was evaluated as the ratio of immunohistochemically stained area for MCP-1 and p47-phox to WGA (%). Three glomeruli were randomly selected in each mouse in all groups, and the mean of all glomeruli selected in each group (i.e., n ×3) was defined as MCP-1 or p47- phox staining area/WGA.
Images under microscopy were scanned into a computer using dedicated software (Olympus DP-controller DP manager®, Olympus). Calculation of ECM, MCP-1 or p47-phox stained area, and WGA was performed using Win ROOF Ver 5.8.1® (Mitani Co., Ltd., Fukui, Japan).
Quantitative analysis of mRNAs in renal cortex by real-time polymerase chain reaction
All probes and primers used for quantitative real-time PCR assay were purchased from Applied Biosystems (Applied Biosystems, Co., Ltd., Foster City, CA, USA) (TaqMan® Gene Expression Assays). Total RNA was extracted using ISOGEN® (Nippon Gene, Co., Ltd., Toyama, Japan) and was converted to cDNA using an ExScript RT Reagent Kit® (Takarabio, Co., Ltd., Shiga, Japan) with random oligonucleotide primers. Real-time PCR was performed using an Applied Biosystems 7500 (Applied Biosystems) with TaqMan® Universal PCR Mix (Applied Biosystems) and gene-specific primers. PCR conditions were 50°C for 2 min and 95°C for 10 min, followed by 50 cycles of 95°C for 15 s and 60°C for 1 min. GADPH mRNA (Applied Biosystems) was used as a control for RNA template normalization. Relative quantitation was performed using the comparative CT method, and data are given as a ratio against the control C57BL/6J mice group.
Statistical methods
All data are presented as means ± standard deviation (SD). Multiple comparisons among more than 3 groups were performed using analysis of variance (ANOVA). If ANOVA was significant, a post-hoc Bonferroni test was used. P values of less than 0.05 were considered to indicate statistical significance in all analyses.
RESULTS
BW, BP and blood and urine test data
Two mice in the CA, and 1 mouse in the CDCA groups died before 22 weeks of age; therefore, the number of mice at 22 weeks was 6 in the CA group, 7 in the CDCA group. Casual BG in the 3 KKAy groups was higher than in the C57BL/6J group at 14 and 18 weeks of age. At 22 weeks of age, casual BG in the KKAy control and CDCA groups was higher than in the C57BL/6J group. There were no significant differences in casual BG between the CA and C57BL/6J groups, and casual BG in CA (but not CDCA) was significantly lower than in KKAy control mice. Significant increases in BW in the 3 KKAy mice group were noted when compared with the C57BL/6J group, while no significant differences were observed among the 3 KKAy mice groups at 14, 18 and 22 weeks of age. With regard to blood pressure, no significant differences were observed among the 3 KKAy groups and C57BL/6J mice, except at 14 weeks of age between C57BL/6J mice and the KKAy control and CA groups, and for DBP between C57BL/6J mice and CA mice at 22 weeks of age. Casual BG, BW, SBP and DBP data in the 3 KKAy groups and C57BL/6J mice at each observation point are shown in (Figure 1A-1D).
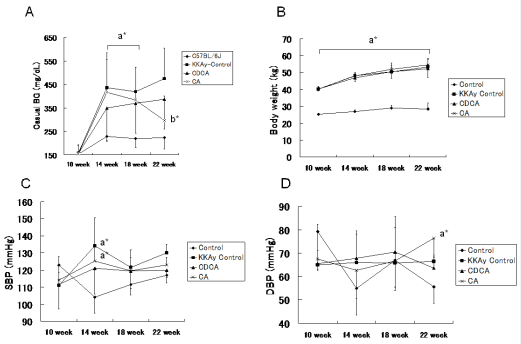
Figure 1 A) Casual blood glucose (BG) at 10, 14, 18 and 22 weeks of age in C56BL/6J (control) mice, KKAy mice without drugs (KKAy control), and KKAy mice treated with chenodeoxycolic acid (CDCA) or colic acid (CA). a* indicates significant difference between C57BL/6J mice and KKAy control, CDCA, or CA mice at 14 and 18 weeks of age. b*, c* indicates significant difference vs. control (C56BL/6J) and KKAy control mice, respectively. Data are shown as means ± SD. B) Body weight (BW) at 10, 14, 18 and 22 weeks of age in C56BL/6J (control) mice, KKAy mice without drugs (KKAy control), and KKAy mice treated with chenodeoxycolic acid (CDCA) or colic acid (CA). a* indicates significant difference between C57BL/6J mice and KKAy control, CDCA, or CA mice at 10, 14, 18 and 22 weeks of age. C) Systolic blood pressure at 10, 14, 18 and 22 weeks of age in C56BL/6J (control) mice, KKAy mice without drugs (KKAy control), and KKAy mice treated with chenodeoxycolic acid (CDCA) or colic acid (CA) a* indicates significant difference from C57BL/6J mice at 14 weeks of age. D) Diastolic blood pressure at 10, 14, 18 and 22 weeks of age in C56BL/6J (control) mice, KKAy mice without drugs (KKAy control), and KKAy mice treated with chenodeoxycolic acid (CDCA) or colic acid (CA) a* indicates significant difference from C57BL/6J mice at 22 weeks of age.
With regard to serum lipids at 22 weeks of age, significantly elevated TG, HDL-C and LDL-C in KKAy control mice was noted as compared with C57BL/6J mice. There was a significant decrease in LDL-C, and a tendency for decreased HDL-C in the CDCA group, and a significant decrease in TG in the CA group, as compared with KKAy control mice. With regard to TBA, significant increases in CA-treated mice were observed, as compared with C57BL/6J and KKAy control mice at 22 weeks of age. Significant increases in UAE in the KKAy control, CDCA and CA groups were observed, as compared with C57BL/6J mice. Both CDCA and CA showed significantly reduced UAE, as compared with KKAy control mice. Data on blood and urine tests in C57BL/6J and the 3 KKAy groups at 22 weeks of age are shown in (Figure 2).
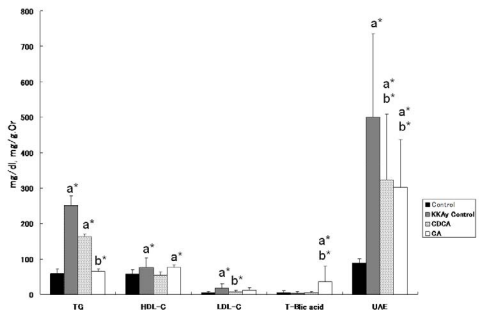
Figure 2 Blood and urine findings at 22 weeks of age in C56BL/6J (control) mice, KKAy mice without drugs (KKAy control), and KKAy mice treated with CDCA or CA. BG: blood glucose; TC: total cholesterol; TG: triglyceride; HDL-C: high density lipoprotein cholesterol; LDL-C: low density lipoprotein cholesterol; TBA: total bile acid; UAE: urinary albumin excretion. a*, b* indicates significant difference vs. control (C56BL/6J) and KKAy control mice, respectively. Data are shown as means ± SD.
Light microscopy and immunohistochemical staining
On real-time PCR of the renal cortex, significantly elevated mRNA expression of MCP-1, fibronectin, nicotinamide adenine dinucleotide phosphate (NADPH) oxidase 4 (NOX4), farnesoid X receptor (FXR) and stimulating sterol regulatory element-binding protein 1c (SREBP1c) in KKAy mice was observed, as compared with C57BL/6J control mice. MCP-1, fibronectin and SREBP1c were significantly decreased in the CDCA and CA groups, as compared with KKAy control mice. These data are shown in (Figure 3).
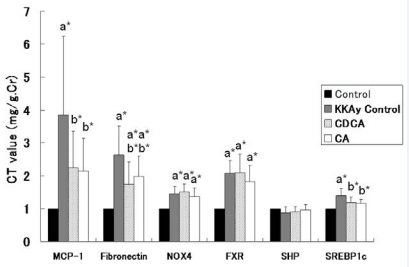
Figure 3 mRNA levels in renal cortex at 22 weeks of age in C56BL/6J (control) mice, KKAy mice without drugs (KKAy control), and KKAy mice treated with CDCA or CA. mRNA levels were evaluated by comparative CT method, and are expressed as ratio vs. C56BL/6J (control) mice MCP-1: monocyte chemoattractant protein-1; NOX4: nicotinamide adenine dinucleotide phosphate oxidase 4; FXR: farnesoid X receptor; SHP: small hetelodimer partner; SREBP1c: stimulating sterol regulatory element-binding protein 1c. a*, b* indicates significant difference vs. control (C56BL/6J) and KKAy control mice, respectively. Data are shown as means ± SD.
On PAS staining of renal cortex, as evaluated by light microscopy, marked increases in mesangium matrix and cells in glomeruli were evident in KKAy control mice when compared with C57BL/6J mice. CDCA and CA inhibited the extent of increase of mesangium matrix and cells. There was a significant increase in ECMA/WGA in KKAy control mice, as compared with C57BL/6J mice. CDCA and CA showed significantly reduced ECMA/ WGA when compared with KKAy control mice (Figure 4A). On immunohistological staining for MCP-1 and p47-phox, increased MCP-1 and p47phox staining in glomeruli was observed in KKAy control mice, as compared with C57BL/6J mice. CDCA and CA showed significantly decreased MCP-1 and p47-phox staining area/WGA when compared with KKAy control mice (Figure 4B,4C).
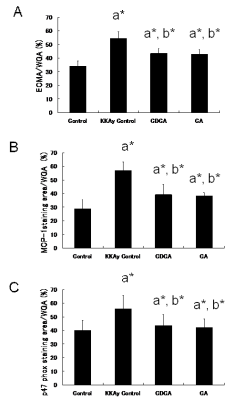
Figure 4 ECMA/WGA ratio at 22 weeks of age in renal cortex stained with PAS (A), MCP-1 staining area/WGA ratio at 22 weeks of age in renal cortex with immunohistochemical staining for MCP-1 (B), and p47 phox staining area/WGA ratio at 22 weeks of age in renal cortex with immunohistochemical staining for p47 phox (C). ECMA: PAS-stained extracellular mesangial matrix area; WGA: whole glomerular area. Three glomeruli were randomly selected in each mouse, and evaluation was performed for all mice in each group (i.e., n×3). a*, b* indicates significant differences vs. control (C56BL/6J) and KKAy control mice, respectively (P<0.001).
DISCUSSION
In the present study, CA significantly decreased casual BS levels at 22 weeks of age, as compared with KKAy control mice, thus supporting the results of previous studies [2,3]. On the other hand, CDCA did not significantly affect casual BG, although a slight decrease (not significant) was noted. It is difficult to explain why CA alone showed glucose-lowering effects, and only CA significantly increased serum TBA levels (probably CA; we could not investigate BA isoforms in this study). It has been reported that the TGR agonist effects of CDCA and CA are similar [4]; therefore, it is possible that marked increases in TBA by CA administration contribute to the improvement of glycemic control via the TGR5-cAMP-D2 pathway in skeletal muscle. On the other hand, in this study, CDCA and CA similarly decreased mRNA expression of SREBP1c, which is associated with synthesis of fatty acid [12], as compared with KKAy control mice.
As the SREBP1c gene is thought to be located downstream of the FXR gene, the effects of CDCA and CA on renal FXR may have been equal in this study. Although FXR activity is generally stronger with CDCA than with CA, as reported previously,[1] it is also possible that CDCA does not exhibit its full effects on FXR in liver because of relative small increases in CDCA in circulation; we speculate that although CDCA scarcely affected circulating TBA levels, the ratio of CDCA in TBA in circulation was elevated. Furthermore, some studies have failed to demonstrate the glucose-lowering effects of GW4064 (selective FXR agonist) [2,13], which does not have a TGR5 agonist effect. This suggests that the glucose-lowering effects that occur via the FXR-PEPCK pathway of BA are mild when compared with those via the TGR-cAMP-D2 pathway of BA. Taken together, we speculate that CA has a stronger glucose-lowing effect because CA can more effectively elevate circulating TBA levels when compared with CDCA, resulting in improved glucose metabolism via the TGR5- cAMP-D2 pathway.
Notably, despite the differences in the effects of CDCA and CA on glucose levels, the decrease in UAE was similar. Furthermore, CDCA and CA significantly decreased mRNA expression of MCP-1, which is associated with renal inflammation [14,15], and fibronectin in the renal cortex, as compared with KKAy control mice, in addition to the improvement in microscopic histological findings evaluated by PAS staining, and MCP-1, p47 phox (a component of NADPH oxidase) immunohistochemical staining. These results suggest that CDCA and CA can inhibit the progression of diabetic nephropathy to the same degree in vivo. We speculate that the effects of CDCA on diabetic nephropathy are based on its strong FXR agonist action independent of glycemic control. On the other hand, the effects of CA appear to be more complex, and its glucose-lowering effects may also be important for improving diabetic nephropathy.
With regard to TG, CA effectively decreased TG levels when compared with CDCA. Although SREBP1c mRNA in the liver was not investigated in this study, as renal SREBP1c mRNA expression in the CDCA and CA groups was similar to previous reports, we speculate that the decrease in TG in CA mice group is based on improved insulin resistance by remission of glucose toxicity.
In conclusion, administration of CA, but not CDCA, resulted in marked elevation of TBA in KKAy mice, and only CA significantly decreased casual BS levels when compared with KKAy control mice at 22 weeks of age. Nonetheless, both CDCA and CA inhibited the progression of diabetic nephropathy. These results suggest that CDCA can improve diabetic nephropathy independently of glycemic control, probably via FXR in the kidney because of its potentially strong effects on FXR, and that the glucose-lowering effects of CA are more complex. Additional studies into whether higher dose CDCA, which may increase circulating TBA levels, can provide more beneficial effects for diabetic nephropathy and glycemic control are necessary.
ACKNOWLEDGEMENT
We are grateful to that staff at the Joint Research Center and of Pathological Division of Dokkyo Medical University Koshigaya Hospital.









































































































































